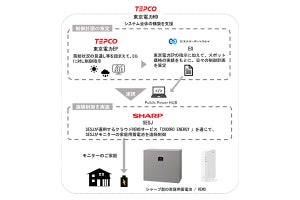
理化学研究所(理研)は5月8日、筑波大学との共同研究により、環形動物のゴカイの体節形成を詳細に観察し、従来の説とは異なり、新たな体節は隣接する手前の体節からのタンパク質が増殖シグナルとなって作られる仕組みであることを発見したと発表した。
成果は、理研 発生・再生科学総合研究センター 形態形成シグナル研究グループの丹羽尚研究員(現・客員研究員)、同・秋元愛テクニカルスタッフ(現・自然科学研究機構 基礎生物学研究所 IBBPセンター)、同・林茂生グループディレクター、同・ゲノム資源解析ユニットの工樂樹洋ユニットリーダー、筑波大大学院 生命環境科学研究科の佐久間将研究員らの共同研究チームによるもの。研究の詳細な内容は、米科学誌「Developmental Biology」オンライン版に近日中に掲載される予定だ。
ヒトの脊椎骨や蛇の胴体のように、生物の体には構造単位が繰り返される部位がよく見られる。この繰り返される単位は「体節」と呼ばれ、脊椎動物だけでなく、昆虫類や甲殻類を含む節足動物などにも見られる生物の形態デザインの基本単位だ。
脊椎動物や節足動物では、例えばショウジョウバエは14個、ヒトは30個と、体節の数はあらかじめ決まっている。これらの動物の発生過程では、胚が伸長していく最先端部に大規模な増殖領域が生じ、そこから細胞が供給されて体節が作られる仕組みだ。増殖領域は、発生過程が終了し個体が形成されると失われるために、ある種のトカゲやイモリといった例外を除けば、成体には体節再生能力はない。
しかし、体節が連なった構造がとてもわかりやすい動物たちである、ミミズやゴカイ、ヒルなどの「環形動物」は異なる。環形動物とは、体が細長く左右対称で目立つ付属肢(手足)がない上に、体に骨格がなくて環状で、体節が直列に並んだ構造を持つ動物たちだ。
この環形動物の内のゴカイを例に取ると、その発生過程では胴部と尾部の間に体節を繰り返し付加し続けて成長する。ゴカイの体節は「体壁」、付属肢、筋肉、消化管、筋肉をすべて備えた円筒状の構造をしており、成体では頭部と尾部の間に大編成の貨物列車のように連結され、その数は120~130にも達するという(画像1)。
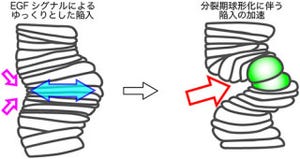
しかも、ゴカイは体を切断されると傷を修復し、体節形成(増節)を加速させて再生するため、ヒトなどとは異なり、実質上無限の体節再生能力を持つと考えられている。研究チームは、この強靭なゴカイの再生能力の謎を探るため、増節の仕組みの解明に挑んだというわけだ。

|

|
|
画像1。イソゴカイの成体。多数の体節に付属肢を持つ |
画像2。イソゴカイの走査電子顕微鏡写真 |
研究チームは、まずゴカイの中でも釣りエサとしてなじみ深い、海岸部に生息する多毛綱ゴカイ類の「イソゴカイ」を用いて、増節の仕組みを詳しく観察することにした。通常、実験室の飼育条件下で成長するゴカイは、およそ4日に1体節の割合で最後端の体節と尾部の境界から増節するという。
増節の時は細胞増殖が盛んになるので、DNA合成期の細胞を可視化する染色法を用いて、細胞増殖の様子、特に体壁を構成する「外胚葉」(初期胚を構成する胚葉の1種で、で、これに属する細胞群は体の外部を取り囲み、表皮とそれに付随した付属肢、神経系などに分化する)での細胞増殖と体節の形成についての観察が詳しく行われた。その結果、新たな体節が付加される時は、最後端の体節の中でも尾側にある1列の細胞群で増殖が活発になることが見出されたのである。
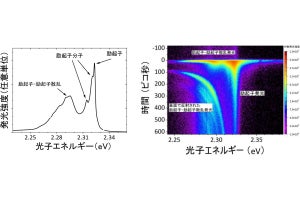
次に、尾部切断後の再生時における増節の調査が行われた。すると、まず切断部に増殖領域が生じて尾部の再生が始まることがわかった。それから数日経って尾部の形成が完了し、その後に増殖領域は切断された体節の尾側に限局したのである(画像3・B'の矢印および画像4)。そして、通常時と同様に1列ごとの規則正しい細胞増殖を起こし、1日に1体節の割合で増節したという。つまり、再生時には通常時の約4倍の速さで増節した計算になる。
こうして列ごとに細胞増殖が活性化し、腹側で細胞が5列に並ぶと1体節の原型ができ上がる(画像4)。その後、細胞増殖がさらに盛んになり、神経、筋肉、内蔵などの内部器官や付属肢が加わり、1つの体節として成熟していく仕組みなのがわかった。
これらの結果から、ゴカイの体節形成は、発生期にある脊椎動物や節足動物の多くの胚で見られるように、伸長の先端にある大規模な増殖領域から細胞が取り分けられるのではなく、最後端の体節の尾側という極めて局所で起こる規則的な細胞増殖に起因することが判明したというわけだ。
さらに研究チームは、この規則正しい細胞増殖の分子メカニズムの解明に取り組むことにした。脊椎動物や節足動物の増殖領域には、細胞増殖と細胞分化を決定するためのシグナル分子としてさまざまなタンパク質が働いている。その代表的なものが「Wnt(WntWingless:ウィント)」だ。Wntは増殖領域で増節に必須の働きをする。
そしてゴカイでWntと同様な機能を持つのが「Wingless(Wg)」だ。Wgは、ショウジョウバエの変異体の解析により、羽がない(Wingless)表現型から見出された分泌性のタンパク質で、その後、ほ乳類でも同様の分子Wntが発見された。WgまたはWntが分泌されると、近傍の細胞膜上にある受容体に結合して細胞の増殖や分化が促される。
WgやWntは胚発生や発がんに深く関与しており、初期発生においては体軸の決定、体節形成、四肢や羽の形成などさまざまな場面で重要な役割を果たす。現在までにWgやWntと類似した分子が19種以上同定されており、これらは総称してWntファミリーと呼ばれている。
そのWgの様子を調べると、既存の各体節の尾側で列となって発現していたが、尾部ではほとんど発現していないことが判明した(画像3・A、B、画像4)。また、最後端の体節にある増殖細胞の列は、既存の各体節にあるWgの発現と平行して起こり、5列に達して増節が完了すると新たなWgが発現して、次の増節サイクルが始まることがわかった。この結果から、最後端の体節からWgタンパク質が増節中の体節内に伝搬し、細胞増殖を規定するという仮説が提示されたのである(画像4)。
画像4を捕捉すると、体節は3→2→1→0と順次形成される。WgとHhタンパク質は、体節境界を挟む1列の細胞でそれぞれ発現する。0番の未成熟な体節では、尾部側の1列の細胞で増殖が活性化する(画像3・BおよびB'の矢印)。1列ごとに細胞増殖が活性化し、細胞が5列に並ぶと1体節の原型ができ上がる。細胞増殖は1番体節でのWgの誘導を受けて起きると考えられるという。

|

|
|
再生中のゴカイ後端部での体節形成の様子とモデル。画像3(左)のAおよびBは、シグナルタンパク質Wg(赤)とHh(黒)の発現パターン。同じくA'およびB'は、細胞増殖マーカー(PCNA、緑)とシグナルタンパク質(Hh、黒)の発現パターン。画像4は、ゴカイの増節モデル |
|
この仮説を検証するために研究チームは、Wgの作用を増強させる「塩化リチウム(LiCl)」溶液中でゴカイの飼育を実施。すると、通常5列からなる1体節の幅が拡大し、1つの体節を形成する速度が遅くなった。この結果は、隣接した体節由来のWgタンパク質の量と伝搬する範囲が、新たな体節の位置とサイズを決定するというモデルを示唆するとする。
今回の研究により、ゴカイの体節形成は、従来知られていた「伸長の最先端部に形成された"増殖領域"で合成されるWgタンパク質が新たな体節形成を促進する」という仕組みではなく、「"既存の体節"から供給されるWgタンパク質が細胞増殖を制御して、新たな体節を形成させる」ということが判明した。研究チームによれば、この仕組みにより、実質上無限に増え続けることができるゴカイの体節の形成能力を説明できるという。
1927年にドイツの実験発生学者マンゴルドとシュペーマンは、両生類胚に移植した神経組織が周囲の細胞に働きかけて神経形成を誘導することを発見し、「相同形質誘導(Homeogenetic Induction)」と名付けた。相同形質誘導はすでに分化した組織・細胞からの誘導によって、近接する未分化な細胞が相同な形質を獲得する現象のことで、体節のように相同の形質が繰り返された構造体を形成するには合理的なメカニズムである。
今回ゴカイで見出された増節の仕組みは、その相同形質誘導が再生の場面で用いられているとする最初の報告だと考えられるという。そして研究チームは今後の課題として、Wgが作用する仕組み、次の体節形成に移行する仕組みなどを明らかにすることとしている。