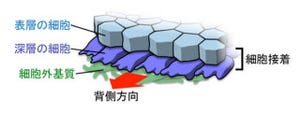
東北大学は、発生期の哺乳類神経幹細胞において、細胞周期調節因子「Cyclin D2(サイクリン D2)」が「脳原基」の外側である「基底膜面」の先端の「基底膜面突起」(画像1)に局在することを発見し、神経幹細胞が2つの娘細胞に分裂する際に基底膜面突起と共にCyclin D2は片方の娘細胞にのみ受け継がれ(残りはニューロンとなる)、その細胞の運命を未分化な状態に維持する働きがあることを解明した。成果は、東北大学大学院医学系研究科の大隅典子教授、恒川雄二研究員(現Scripps研究所所属)らによるもので、詳細な内容は欧州科学誌「EMBO Journal」に掲載される予定だ。
大脳発生初期において神経幹細胞は対称分裂を繰り返し、未分化な細胞を増殖させていく仕組みを持つ。必要な数の神経幹細胞が産生され、ニューロン産生期に入ると分裂パターンに変化が生じ、非対称に分裂するものが現れ始めるのが特徴だ。
片方の娘細胞は細胞周期を逸脱してニューロンへと分化し、一方基底膜面突起を受け継いだもう片方の細胞は再び細胞周期に戻ることにより、未分化性を維持した神経幹細胞となるのである(画像1)。
画像1は大脳発生の模式図であり、増殖期において神経幹細胞が対称分裂で神経幹細胞の数を増やした後、ニューロン産生期になって、1つの神経幹細胞と1つの分化したニューロンを生み出す非対称分裂を繰り返すようになる。この分裂様式により、生み出されるニューロンの数がコントロールされ複雑な大脳が形成されていくのだ。

|
|
画像1。大脳発生の模式図 |
しかし、哺乳類神経幹細胞の非対称分裂において、娘細胞に非対称性を生じさせる運命決定因子、またその非対称分配のメカニズムの多くは未知のままだったのである。
研究グループは、非対称分裂の際、基底膜面突起が片方の娘細胞のみに受け継がれる現象に着目し、細胞周期調節因子として知られているCyclin D2のmRNA(メッセンジャーRNA)およびタンパク質が、ニューロン産生期において神経幹細胞の基底膜面突起に局在していることを発見した。それを撮影したのが画像2で、E(胎生)10.5、12.5、14.5日マウスの終脳におけるCyclin D2のmRNAおよびタンパク質の発現パターンである。
画像2の(A-C)は、E10.5、12.5、14.5日マウス終脳でin situ hybridization(ISH)法を用いてCyclin D2 mRNAの発現パターンを解析したもの。Cyclin D2 mRNAはすべての発生期において「VZ(脳質帯)」と呼ばれる神経幹細胞の存在する層と、神経幹細胞基底膜面突起が存在する基底膜面に存在することが示された(矢頭)。
下段の(D-I)は、E10.5、12.5、14.5日マウス大脳皮質におけるCyclin D2のmRNAおよびタンパク質の発現をISH法および抗体染色法により解析したものだ。E10.5(増殖期)においてはCyclin D2 mRNAは基底膜側突起先端に存在するが、タンパク質は核にのみ存在し、細胞間での発現レベルに差はみられない(E 拡大図)。
E12.5、E14.5(ニューロン産生期)になるとCyclin D2タンパク質はmRNAと同様に基底膜側突起先端に存在するようになり、核においても細胞間で発現レベルの差が確認できるようになっている(G:拡大図の矢および二重矢)。

|
|
画像2。E(胎生)10.5、12.5、14.5日マウスの終脳におけるCyclin D2のmRNAおよびタンパク質の発現パターン |
これらの結果から、研究グループはCyclin D2が神経幹細胞の非対称性に関わっているという仮説を立てこれを証明する実験を始めたというわけである。
研究グループは最初、Cyclin D2 mRNAには細胞体から基底膜面突起に移動するのに必要十分な、輸送配列が存在することを明らかにした。このような配列は世界で初めて報告されたものである。さらに、胎仔脳に対する「電気穿孔法」および全胚培養法により、Cyclin D2 mRNAが輸送された先の基底膜面突起で局所的に翻訳されていることが確認された。
次に、レンチウイルスを用いて非対称分裂後の姉妹細胞を可視化し、基底膜面突起を受け継いだ娘細胞が有意にCycin D2を発現し、その細胞の運命が幹細胞であることも解明している(画像3)。
画像3が、そのレンチウイルスを用いてE14.5の神経幹細胞を可視化し、分裂直後であるG1期の細胞においてのCyclin D2の局在を観察したもの。(A-C)は、Cyclin D2はG1期初期から基底膜面突起を受け継いだ娘細胞(C')に有意に発現が強く、受け継がない細胞では弱く(C")、G1期後期にはその差が顕著に観察された(D-F")。さらに、Cyclin D2は神経幹細胞基底膜面突起先端にも局在していることが観察されたのである(A-F・右上拡大図)。
Cyclin D2を細胞に強制発現させると幹細胞として振る舞い、逆にsiRNAによるCyclin D2の機能欠失を行うと分化したニューロンになることが判明したことから、Cyclin D2が細胞の運命を未分化な状態に維持している働きがあることが判明した(画像4)。
画像4は、Cyclin D2は神経幹細胞の運命決定に寄与しているのを写真とグラフでまとめたものだ。(A-D)はE12.5日マウス胎児大脳皮質に電気穿孔法を用いてコントロール空ベクター(control plasmid)、Cyclin D2発現ベクター(Cyclin D2 ORF)、コントロールsiRNA(control siRNA)、Cyclin D2に対するsiRNA(si1726)をEGFP発現ベクターと共に導入した。
48時間後の大脳切片を成熟したニューロンが存在する皮質板(CP)、移動中の未熟なニューロンが存在する中間層(IZ)、神経幹細胞が存在する脳質下帯(SVZ)、VZに分け、EGFP陽性細胞の局在を観察したものである。
(E)のグラフは、観察の結果Cyclin D2が過剰に存在すると優位に神経幹細胞の割合が増えてニューロンの割合が減り、またその逆の現象がCyclin D2の機能欠失を行った場合に観察されたのをまとめたもの。これらの結果から、Cyclin D2が神経幹細胞の運命決定に寄与していることが示唆された。

|

|
|
画像3。レンチウイルスを用いてE14.5の神経幹細胞を可視化し、分裂直後であるG1期の細胞においてのCyclin D2の局在を観察したもの |
画像4。Cyclin D2は神経幹細胞の運命決定に寄与しているのを写真とグラフでまとめたもの |
一連の結果から想定されたのが、次のようなメカニズムだ。まず、Cyclin D2 mRNAは特異的な配列を基に基底膜面突起に輸送され、局所的に翻訳されることにより基底膜面突起に蓄積される。非対称分裂の際に片方の娘細胞に基底膜面突起が受け継がれることにより、そこに蓄積していたCyclin D2が非対称に片方の娘細胞に受け継がれ、その細胞を未分化な状態に維持。このことにより分裂した姉妹細胞間において運命の非対称性が生まれるというわけだ(画像5)。
画像5は、前述したCyclin D2の神経幹細胞における非対称運命決定機構の仮説の模式図である。Cyclin D2を受け継いだ細胞は自己増殖を繰り返す幹細胞の1つとして増殖を繰り返し、受け継がなかった娘細胞は分化したニューロンとなるというわけだ。

|
|
画像5。Cyclin D2の神経幹細胞における非対称運命決定機構の仮説の模式図 |
Cyclin D2は胎生期のヒト終脳においても基底膜面突起に局在し、輸送配列が存在した(この配列はチンパンジー、アカゲザル、オラウータンでも72~80%で保存されている)。なお、大脳皮質に層構造を持たないニワトリでは、Cyclin D2の局在パターンも輸送配列も確認されておらず、研究グループではその点について興味深いこととしている。
これらの結果は、Cyclin D2の非対称分裂時における未分化性維持が進化の過程で哺乳類が獲得したメカニズムである可能性を示唆するものだ。また、今回の研究成果から、胎生期神経幹細胞において、幹細胞を維持することにおいて基底膜側の突起の重要であることが強く示唆された。
このことは、ヒトを含む霊長類の大脳皮質原基において、基底膜側突起を有する神経幹細胞が多数存在することに鑑みて、多数のニューロンを産生して大きな大脳皮質を構築するストラテジーの一端を示していると思われると、研究グループではコメントしている。