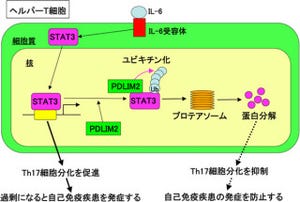
理化学研究所(理研)は、動物細胞の細胞内小器官の中に古細菌型の脂質が存在することを明らかにし、古来の動物細胞が古細菌と接点を持っていた可能性を示唆した。同成果は、理研基幹研究所 小林脂質生物学研究室のPeter Greimel研究員、Hui-Hui Tan国際プログラムアソシエイト、牧野麻美研究員および小林俊秀主任研究員らの研究チームによるもので、独学術雑誌「Angewandte Chemie International Edition」(オンライン版)に掲載された。
すべての生物の細胞膜は、グリセロールにリン酸が結合したグリセロリン酸に、2分子の疎水性の高い分子(脂肪酸など)が結合した脂質(グリセロリン脂質)の二重層でできている。天然のグリセロリン酸には、リン酸の結合部位の違いにより、ちょうど鏡に写したような関係にあるsn-1型とsn-3型の2つが存在している。

|
|
図1 2つのグリセロリン酸の関係。細胞膜脂質を構成するグリセロリン酸にはsn-1型(図中右)とsn-3型(図中左)の2つがあり、これらは左手と右手の関係のように鏡に写した関係にある。赤は酸素原子、黄色はリン原子、青は炭素原子、白は水素原子を表す |
この2つの構造は非常に似ているように見えるが、実際はまったく異なる分子で、哺乳類の動物細胞をはじめとした真核生物からバクテリアと呼ばれる真正細菌(細菌)まで、細胞膜の形成のためにsn-3型だけを選択的に合成して利用することが知られている。一方、生物には真核生物と真正細菌の他に古細菌と呼ばれるグループが存在しており、この古細菌は、細胞膜の形成のためにsn-1型だけを選択的に合成して利用する。グリセロリン酸に疎水分子を結合することは、脂質の合成の最も初期の反応であることから、古細菌は進化の過程の非常に早い時期に、他の生物とはまったく異なる進化をたどったと考えられている。
真核生物は、エンドサイトーシスというメカニズムにより、異物や栄養を細胞膜で取り囲み、小胞を形成して細胞内に取り込む。さらにこの小胞が細胞内小器官(初期エンドソーム)と融合し、そこから再び小胞が形成されて次の細胞内小器官である後期エンドソームに受け渡すことを繰り返す。後期エンドソームは、細胞が取り込んだタンパク質やsn-3型の脂質などを分解するが、その際、ビス(モノアシルグリセロ)リン酸(BMP)と呼ばれる脂質がその他の脂質の分解を促進する。このBMPが分解されずに機能を発揮できる理由として、その構造が他の脂質と異なるsn-1型だからではないかと示唆されているが、その確証は得られていなかった。

|
|
図2 エンドサイトーシス経路と後期エンドソーム。 |
BMPは、脂質の基本構造であるグリセロリン酸にグリセリンが結合したジグリセロリン酸に、2分子の脂肪酸が結合している。

|
|
図3 ビス(モノアシルグリセロ)リン酸(BMP)。ベビーハムスター腎臓細胞では全リン脂質のうちBMPが占める割合は1%以下。BMPはグリセロリン酸にさらにグリセリンが結合した骨格構造に2分子の脂肪酸が結合している。今回の研究でこのグリセロリン酸がsn-1型であることが明らかになった |
研究チームは核磁気共鳴(NMR)を用いて分光学的にジグリセロリン酸部分の立体構造を解析した。実際のNMRによる立体構造の解析には、適当な分子でジグリセロリン酸を修飾し、NMRで判別できる形にする必要があるため、研究チームはsn-1型であるsn-1,1'-ジグリセロリン酸を化学合成した後、修飾剤であるD-カンファー(樟脳)を用いて、化合物(D-カンファービスケタール誘導体)を合成した。また、同様に、理論的に存在可能な立体配置であるsn-3型のsn-3,3'-ジグリセロリン酸、およびsn-3,1'-ジグリセロリン酸のD-カンファービスケタール誘導体も合成した。D-カンファーは、特定の方向に振動する光(偏光)があたると、その面が回転する性質(光学活性)を持つため、このようにD-カンファーで修飾すると、NMRスペクトルの位置関係から、グリセロリン酸がsn-1型なのかsn-3型なのか解析することができるという特長を持っている。
一方、実際の哺乳類動物の培養細胞であるベビーハムスター腎臓細胞から、細胞中にあるリン脂質の1%以下しかないBMPを精製し、不要な脂肪酸部分を除去してジグリセロリン酸のD-カンファービスケタール誘導体を合成した。

|
|
図4 D-カンファーケタール。D-カンファー(樟脳)は立体構造解析で良く使われる修飾剤。ジグリセロリン酸骨格に結合したD-カンファーケタールの位置関係に基づく、電子状態の違いに起因したプロトン(1H)の共鳴周波数のずれ(NMRの化学シフト)からBMPの構造が明らかになった |
このNMRスペクトルを測定し、合成した化合物の測定結果と比較した結果、哺乳類の細胞のBMPから調製したジグリセロリン酸由来のスペクトルは、化学合成したsn-1,1'-ジグリセロリン酸由来のスペクトルと一致することが見いだされ、このことから、哺乳類の細胞にsn-1型の脂質が存在することが明らかとなった。

|
|
図5 動物細胞由来BMPのジグリセロリン酸骨格の立体配置の同定。対照化合物のプロトンNMRスペクトル(A~C)および動物細胞BMP誘導体(D)。Dのスペクトルは、点線で示した部分でAと良く一致し、動物細胞のBMPはsn-1,1'-の構造を持つことが明らかになった |
今回の研究では、真核生物のうち、哺乳類の細胞内のBMPがsn-1型の脂質であることが示されたが、酵母や線虫といった原始的な真核生物からはBMPが検出されておらず、真正細菌ではBMPの報告はあるもののsn-3型であり、動物細胞の後期エンドソームだけにsn-1型の脂質が存在する理由については、明らかになっていない。トキソプラズマのように、感染した寄生虫が後期エンドソームの中で生き続ける例が知られているが、BMPも古細菌が感染した名残なのかもしれないと研究チームでは説明している。
BMPは、その合成経路も分解経路も分かっていない謎の脂質であり、研究チームでは今後の研究でBMPの合成酵素を明らかにし、古細菌の酵素と比較することで古細菌と動物細胞との関係が明らかにできれば、古細菌型の構造をもつBMPが動物細胞のなかに存在する謎を解明できるかもしれないとしており、その結果、脂質の構造から生物の進化の道筋が見えてくることも期待できるとしている。