九州大学は8月29日、脊椎動物の未受精卵の分裂停止の仕組みを分子レベルで解明したと発表した。
今回の研究の成果は、米東部時間9月12日に、米Cell Pressが刊行している科学雑誌「Developmental Cell」に掲載される予定だ。
ヒトを初めとする脊椎動物の未受精卵(成熟卵)は、排卵後に第二減数分裂の中期「Meta-II」で分裂を停止し(Meta-II停止)、受精を待つという仕組みを持つ。Meta-IIでの分裂停止は、未受精卵の単為発生(精子なしでの発生)を防ぐために不可欠な仕組みだ。Meta-II停止を起こす物質を「細胞分裂抑制因子(CSF)」と呼び、今回の研究グループの代表である九州大学大学院理学研究院教授の佐方功幸氏らが1989年に「Mos」というタンパク質がCSFの主成分であることを発見。さらに、「Emi2」と呼ばれるタンパク質もCSFの重要な成分であることを突き止めたのも佐方氏らだ。
こうした従来の研究で判明したのは、Mosが「Mos-MAPK経路」(Mos、MEK、MAPK、RSKの4種類のキナーゼからなるタンパク質リン酸化経路)と呼ばれる分子経路を介してEmi2を安定化・活性化させ、Meta-II停止を引き起こすことが1つ。そして、Emi2がタンパク質「Cdk1」によって不安定化・不活性化させられること、さらにMos-MAPK経路がこれに対して拮抗的に働くことで、Emi2を安定化・活性化させることなども判明した。しかし、Cdk1によるEmi2の不安定化・不活性化、およびMos-MAPK経路によるEmi2の安定化・活性化の分子機構は依然として不明だったのである。
そこで今回の実験では、アフリカツメガエルの卵を用いて、まずCdk1によるEmi2の不安定化・不活性化の機構を解析。Cdk1もMosと同じタンパク質リン酸化酵素(キナーゼ)の一種であることから、Cdk1によってリン酸化を受けるEmi2の部位(アミノ酸残基)を複数決定し、それらをリン酸化されないアミノ酸残基に置換した。
すると、このEmi2変異体は未受精卵内で代謝的に非常に安定であり、かつ未受精卵の分裂後期促進因子「APC/C」を強く阻害し、未受精卵をMeta-IIで停止させる活性を持つことも確認。このことから、Cdk1による特定部位のリン酸化によって、Emi2が不安定化・不活性化することが示されたのである。また、Cdk1によるEmi2のリン酸化部位にCdk1自身およびほかのキナーゼ「Plk1」が結合していることも発見された。
続いて、Emi2に結合したCdk1やPlk1がいかにしてEmi2を不安定化・不活性化させるかを解析したところ、まず結合したPlk1がEmi2のほかの部位をリン酸化し、この部位にタンパク質分解関連酵素の一種である「SCGユビキチンリガーゼ」が結合し、Emi2を不安定化させることが判明したのである。
一方のCdk1はEmi2に結合すると、Emi2の末端部位「RL尾部」(APC/Cとの結合に必要な部分)をさらにリン酸化し、Emi2を不活性化させることが確認された。
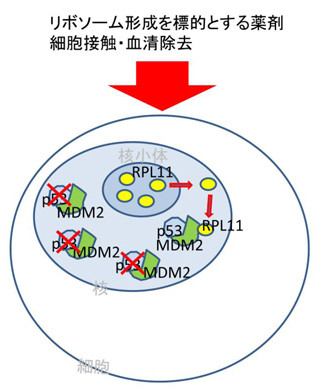
それらに加えて、Cdk1やPlk1の結合部位にはほかのキナーゼの「CK1」も結合し、Cdk1と同様にRL尾部をリン酸化し、Emi2を不活性化させることも判明。まとめると、Cdk1によるリン酸化部位にCdk1自身、Plk1およびCK1が結合し、さらにほかの部位をリン酸化することでEmi2を不安定化・不活性化させるというわけである(画像1)。
そして次に行った解析が、Mos-MAPK経路によるEmi2の安定化・活性化の機構。すると、Mos-MAPK経路によってEmi2がリン酸化され、このリン酸化部位にタンパク質脱リン酸化酵素の一種である「PP2A-B56」が結合することが判明。このPP2A-B56が、Cdk1、Plk1、CK1によってリン酸化した部位を脱リン酸化することで、Emi2を安定化・活性化させるという仕組みである。
結論として、アフリカツメガエル未受精卵内では、Emi2に結合したキナーゼ群によるリン酸化がEmi2に結合したPP2A-B56によって微細に調節されることでEmi2が安定化・活性化され、Meta-II停止が起こることが判明したというわけだ。
今回の結果から、Mos-MAPK経路およびEmi2は脊椎動物未受精卵のMeta-II停止に必須であり、マウスの実験では停止がないと卵巣奇形種が起きてしまうことがわかっている。そこから、ヒトの不妊や卵巣奇形種の原因の1つとして、Mos-MAPK経路の不全やEmi2の変異などの可能性が考えられ、それらの予防・診断や、治療方法開発の足がかりになるとしている。また、受精・再生医学などへの広がりも大いに期待されているとした。

|
|
画像1。Mos-MAPK経路によるEmi2の安定化・活性化の分子機構。最初にCdk1はEmi2の複数の「セリン」(S)、「トレオニン(T)残基」をリン酸化し、これらのリン酸化部位にCdk1自身、Plk1、CK1が結合。結合したPlk1はほかの部位(「DSG」と「DSA」)をリン酸化し、SCFユビキチンリガーゼの結合を介してEmi2を不安定化させる。一方で、Cdk1とCK1はEmi2のRL尾部をリン酸化し、Emi2とAPC/Cの結合を総外資、Emi2を不活性化。これらに対して、Mos-MAPK経路はEmi2をほかの部位でリン酸化し、この部位にPP2A-B56を結合させる。結合したPP2A-B56はPlk1、Cdk1、CK1によってリン酸化された部位を脱リン酸化し、Emi2を安定化・活性化させる。これにより、Emi2はAPC/Cと強く結合し、その活性を抑制することでMeta-IIを引き起こすのである。 |